2023.07.08野菜の抗酸化力を高める「葉っぱ根」で、高品質に安定的多収穫に向けて、生理生態的特性を活性化を推測
青果の抗酸化力を高める「葉っぱ根」とは…
肥料の機能性を活して…野菜の抗酸化力を高めて、高品質に安定的多収穫をサポートします。
「葉っぱ根」は
・土壌の栄養環境を整えます。
植物(野菜)が、健全に生育するための必要な栄養素を根から吸い上げさせる。
・肥料の機能を最大限に発揮させる。
土壌潅水や葉面散布で、自由水を「葉っぱ根」で…さらに代謝水が効率を高めます。
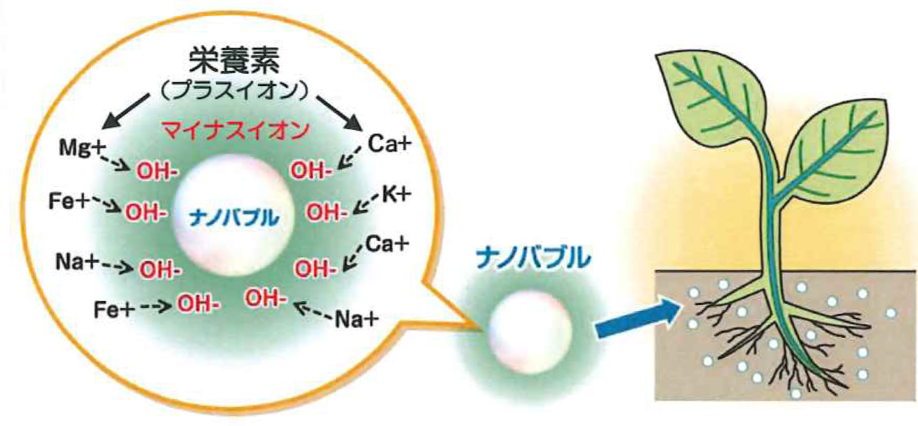
1)水に、ウルトラ ファイン バブルを処理
2)処理水に、微細還元アミノ酸(リズム3)を充填
※自由水のイオン処理で、土壌中の様々な肥料が、代謝水へと変換され…旺盛な生育に。
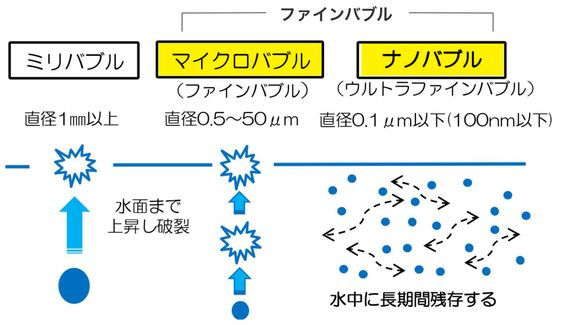
ウルトラ ファイン バブルとは…
ファインバブルは、直径が100μm(=0.1mm)より小さな泡を指す。 (2017年6月にISO規格で定められた)
気泡の大きさが1μm以上の100μm未満はマイクロバブルで、1μm未満はナノバブルとよびます。(別名:ウルトラ ファイン バブルと呼称します)
「葉っぱ根」の使い方
肥料の機能には、即効性, 緩効性, 遅効性といった特徴があります。
植物の種類や生長ステージによって、使い分けることが大切です。
土壌や作物の状態により、使い方や追肥するタイミングを見極めることが重要です。
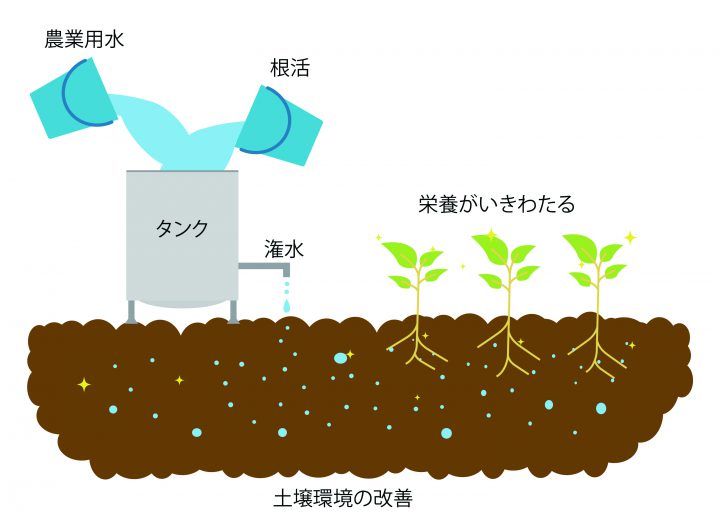
水と空気で出来ているナノバブル水に、微細還元アミノ酸充填溶液は…いろいろな使い方が可能です。
水, 潅水タンク, 養液タンク, 液肥タンクetc…どれに、希釈しても安全です。
イチゴ, トマト, きゅうり, 水稲, 根菜類, 葉菜類, 果樹類, 麦 ,茶葉など、様々な作物の栽培にお使いいただけます。
使用…目安は、灌水時に5000倍以上に希釈して5~10日毎に投与、または2000~5000倍に希釈して5~7日毎で葉面散布する形をおすすめしています。(作物種, 生育ステージ, 栽培環境で異なります)
1回の潅水量は、1反につき原液1~3kg程度で…葉面散布は、葉露程度の量で、多回数でお使いください。
「効果を最大にするために」
- 使用前に下記「注意事項」を確認して下さい。
- 資材を入れる前に、タンクに水を入れて下さい。
- 本製品を良く振ってから、希釈して下さい。
- 本製品を事前に,、適量の水で良く溶いてから、徐々に加えて下さい。
- しっかりと水に溶解することで、資材の効果が最大限に発揮できます。
- タンク内をよく攪拌しながら散布して下さい。
- 使用後は、潅水設備や散布機などの内部を水で良く洗って下さい。
ナノバブルの土壌環境への影響
微細還元アミノ酸充填溶液を散布した区域は、微生物総数が増えたという報告があります。
有機肥料の分解、吸収には微生物の働きが欠かせないため、この点においても微細還元アミノ酸充填溶液の農業利用において有効な効果が得られています。
土着菌や微生物が活性化され増えることは、土壌環境に多くの利点をもたらせます。
肥料の分解/ 溶解作用への貢献は勿論のこと、団粒構造を促し…地温を上昇させ、土壌環境の好環境に繋げることができます。
有機栽培が見直されている現在において、このような土壌改良効果から、今後の「微生物 酵素」に期待がされている分野でもあります。
無駄なく肥料を活かす「葉っぱ根」で、元気に育てる
元肥(もとごえ)
元肥は、苗などを植え付け前に与える肥料を指します。
即効性は期待せずに、長期にわたって植物が順調に育つために、追肥をあたえることです。
効果が長続きする肥料なので、作物によって緩効性や遅効性の肥料を選びましょう。
追肥(ついひ)
追肥は、生育途中で補う肥料で、生育にあわせて不足している栄養を補うことです。
生育途中の植物の状態に対して処置を行いますので、即効性のある肥料が適します。
潅水や葉面散布で行いますが、生育状況によっては、葉面散布が有効です。
寒肥(かんごえ)(冬肥)
寒い時期に与える肥料を「寒肥」と言います。
有機質肥料を使い、微生物や土着菌の働きを活性化します。
冬の間に、土中の成分(残肥)の改善で、植物の生長が活発になる春に栄養素が浸透しやすくなります。
お礼肥(おれいごえ)
花を咲かせた後や、果実を収穫した後に施肥することを指します。
開花や結実に、エネルギー(栄養)を消耗/消化するため、栄養素を補給することで樹勢を回復/強化させます。
芽出し肥(めだしごえ)
芽の生長を促進させる目的で、与える施肥のことを指します。
一般的には、芽が動き始める2月~3月にかけて与えます。
花芽誘導(はなめゆうどう)/ 分化促進
成長点で、葉を形作っていたものが、花を形作るように性質が変化した段階から、花芽が分化したといわれます。
形を変えた葉とは、萼, 花弁, 雄しべ, 雌しべと呼ばれるもので、これらを総称して「花葉」と呼んでいます。
萼, 花弁, 雄しべ, 雌しべは、葉が形を変えた(変態した)ものなので、花が茎の先(成長点)や葉腋(わき芽)につくのは、そのためなのです。
花芽が生育する段階で、複数(花房)の花を作る場合もあります。
トマトなどは、その典型的なもので、花梗(茎)が次々と枝分かれし、その先に花をつけるのです。
これは花梗の先端の芽が先祖返りをして、葉芽になったために発生する現象です。
栽培上は、チッソ過多が原因ということになっていますが、この現象を見ても、花は葉になるべきものが、体内のホルモンの影響で変化したことを示唆しています。
花芽分化と環境条件
炭水化物・・・内生ホルモン・・・植調剤・・・着果負担・・・地温/気温の気象的な推移など、樹体内の栄養条件や栽培環境に起因します。
植物(野菜)が健全に生育する、必要な栄養素を根から吸い上げさせる、土壌の栄養環境を整えてあげることが大切になってきます。
作物の成長を促進させる使い方
土壌環境への影響
抗酸化力の発生について
植物体には、ポリフェノール性物質, フラボノイド, キサントフィルやアスコルビン酸などの種々な化合物が含まれている。
ポリフェノール性物質やフラボノイドについては、病虫害抵抗性やIAAオキシダーゼとの関連, キサントフィルについてはクロロィルによる光エネルギー獲得の共助, アスコルビン酸は、生体の酸化還元をつかさどる物質として知らています。
しかし、近年…生体における活性酸素の発生と、その防御機構が明らかになるにつれ、これらはその重要な機能の一翼を担う物質として、注目されるようになった。
そこで、ここでは活性酸素の種類と生成、活性酸素消去要素による消去および環境と活性酸素の発生について

活性酸素の種類と生成
葉緑体においては通常光エネルギーを捕捉し、PSIでH ,O を H−,Oおよびeに分解し、e-は最終的にPSIにおいてNADPHとATPに変換され,RubiscoによりCOzが同化され,最終的に糖に還元される。
発生したe一がすべてNADPHとATPの生成に使われず、O,と反応し,02-を生成する。
活性酸素消去酵素による、消去および環境と活性酸素
1.スパーオキシド(02 −)葉緑体においては通常光エネルギーを捕捉し, PSl1でH,OをH−, 0 , およびeに分解し、e-は最終的にPSIにおいてNADPHとATPに 変換 さ れ ,Rubisco に よ り COz が 同化 さ れ ,最終的 に糖 に 還元 され る.発生 した e 一が す べ て NADPH と ATP の 生 成 に 使 わ れ ず ,0 , と反応 し ,02一を 生 成 す る .そ の 生 成 速 度 は [5 −30 μ mol /mgCh [/h と さ れ , 光 合 成 電 子 伝 達 速 度 の 5 −10% 以 内 で あ る と い う (高橋 1988 ).強 光 で e一生 成 が 過 剰 な と き,CO2 の 供 給 が NADPH の 消費 を下 回 る と き な ど,こ の 生 成 は増 加 す る (図 1). ミ トコ ン ド リア に お い て も 02 一を発生 す る .発生 さ せ る 系 の 1 つ は NADH 一ユ ビ キ ノ ン レ ダ ク タ ーゼ (複合体 D 系 で NADH が 酸 化 され る と き ,コ エ ン ザ イ ム Q (CoQ ) はCoQ ラ ジ カ ル を生 成 し,こ れ が 0 、と 反応 して ,0 、 一を生 成 す る 。また , ユ ビ キ ノ ール ーシ トク ロ ム c レ ダ ク タ ーゼ 系 (複 合e49111)で も生 成 す る.生 体膜 で は NADPH が NADPH P450 レ ダ ク タ ーゼ ・シ トク ロ ム P −450 系 で 酸 化 さ れ と き,0 、 一を生 成す る (浅出 ら 1992). 2.過 酸化水素 (H202 ) 葉緑 体 に お い て は 0 、 一が ス ーパ ーオ キ シ ド ジ ス ム タ ー ゼ (SOD ) に よ っ て 不 均 化 さ れ て 生 じ る...一方,ペ ロ オ キ シ ソ ーム に お い て は 強光, 高温 な どの と き, リブ ロ ース 2 燐 酸 は Rublsco に よ っ て 酸 化 され (光 呼吸 ), ホ ス ホ グ リ コ ー ル 酸 が 生 じ る.グ リ コ ール 酸が ペ ロ オ キ シ ソ ーム に 移 動 し, グ リオ キ シ ル 酸 に 酸 化 さ れ る と き,H , 02 を生 じる (図 D . 油 脂 を含 む種子 の グ リオ キ シ ソーム で は 発芽 時油脂 が 炭水 化物 に 合 成 さ れ る と き グ リ オ キ シ ル 回 路 で H 、Oz を .生 じ る .ミ ト コ ン ドリア に お い て も生 成 す る.ユ ビ セ ミ キ ノ ン が 0 , を還 元 し,02 −.が生 じ,MnSOD に よ り, H , 0 、 が 生 じ る.ま た , 呼 吸 で 消費 され る 02 の 2% が H20 , とな る (Scun− dalios1994).電子 の 流れ を 阻害す る阻害剤 (パ ラ コ ・一トな どの 除草剤 )は H202 の 発 生 を促進 す る.H /02 は 比 較的安 定 し た 酸化剤 で 反 応 性 が 高い と は い え な い .次 に 述 べ る 活 性 の 高 い ヒ ド ロ キ シ ル ラ ジ カ ル (OH ・) を生 成 す る. 3. ヒ ドロ キ シ ル ラ ジ カ ル (OH ・) 生 体 で は最 も反 応 活 性 の 高 い 活 性 酸 素 で ’site −specific oxidant ’と し て DNA や 膜 脂 質 を 攻 撃 目標 と す る 一方, フ ェ ノ ール 性 化合物 に 水 酸 基 を導 入 す る.葉緑体 に お い て は H 、0 、が 還元 型 フ ェ レ ドキ シ ン と反 応 し, OH ・を つ く る.ま た ,OH ・は Fenton 反 応 (M [1+ + H ,0 、 → M 「u 1 〕1.+ OH ・+ OH .) に よ り生成す る..生 体 で は M は Fe と さ れ , Haber −Weiss 反 応 ; Fe ’3 十 〇∫ → Fe + L 、02 に よ り 2 価 の Fe イ オ ン が 再 生 さ れ , OH ・の 生 成 が 助 長 さ れ る (Scan− dalios i994). 4. 過酸化 脂質 脂 質 の 過酸 化反 応 は 脂質 ア ル キ ル ラ ジ カ ル (L・)の 生 成 か ら 開始 す る.こ れ は 分 子 状 酸素 と 反 応 し ,脂質ペ ル オ キ シ ラ ジ カ ル (LOO ・)を生 成 す る.さ ら に 脂質 か ら 水 素引抜 き ,L・の .生 成 と過酸化脂 質の 生 成 を と もな う.ラ ジ カ ル 連 鎖反応 が 進 み ,過 酸化脂質 の 生 成蓄 積が 起 こ る (脂 質 の 自 動酸化 ).L ・は (,H ・や ア ル コ キ シ ラ ジ カ ル の 作用 で も生 成 す る (寺尾 1988 ).脂質過 酸化反応 の 最終生 産 物 は マ ロ ン ジ ア ル デ ヒ ドと され ,こ の 含 量 を 計 っ て 過 .酸化脂質含 量 と し て い る . 鉢 II.抗酸化 物質 鸞黜 、. 力 。 チ ノ イ ド カ ロ チ ノ イ ドの 機 能 は 集光性 に あ る と考 え られ て き た . 糸[:色 イ オ ウ 細 菌 の カ ロ チ ノ イ ド合成 能 を 欠 い た 変 異 株 は 窒 素中で は 光合成 し,生 長 した が ,空 気 中 で は生 長 が 止 ま り,バ ク テ リオ ク ロ ロ フ ィ ル が 破壊 さ れ た (Sisitrornet al .1957 ). カ ロ チ ノ イ ドを欠 い た トウ モ ロ コ シ は.光酸 素障 害 が 現 れ る (Koshi and Smith l95D 、 ジ メ チ ル ベ ン ズ ア ン ト ラ セ ン に よ る 腫 瘍 発生 遅 延 , 赤 血球 ポ ル フ ィ リ ン 症 発 症 の 遅延 や 癌 発生 と カ ロ チ ノ イ ド含有 野菜摂取量 と逆 相 関 す る こ とか ら もカ ロ チ ノ イ ドは 光酸 素障害 と関係 深 い こ とが 明 ら か とな っ た (牛 島 1988). カ ロ チ ノ イ ドは励 起 1 重項 また は 3 車項 ク ロ ロ フ ィ ル と の 反応 (3Chl 十 1β一caroterle → 1Chl 十 R・β一carotene ),LO2 の 消 去 (102 + 1βcarotene → 30z 43 β .carotene ,た だ し,葉 緑 体 に お け る 1 重 項酸 素 を除 ⇔ お よ び 脂 質 過 酸 化 反 応 の 抑 制 が 知 られ て い る (牛 島 1988).ま た ,葉緑体 で は キ サ ン ト フ ィ ル サ イ クル を形 成 し,過剰 な光 エ ネル ギ ーの 消 去 に 関 与 す る (A ]scher et al.1997). 2. ポ リフ ェ ノ ール マ ク ロ フ ァ ージの ...種 に よ る 活 性 酸 素 の 発 生が タ ン ニ ン 酸 , フ ェ ル ラ 酸 ,コ ーヒー酸 で 抑 制 さ れ る (KhandUja et al . 1997 ).U938 細胞 に よ る O , 一の 生産 は 植物 フ ェ ノ ール で 押 さ え られ る (ク ロ ロ ゲ ン 酸 , コ ーヒ ー酸,ケ ル セ チ ン ,ク マ リ ン , エ ピ カ テ キ ン な ど, F1・oemming und O ’brien 1997). 3. フ ラ ボ ノ イ ド フ ラ ボ ノ ール は 0 ,一,H 、O 、や OH ・を 除 去 す る (高 濱 1988 ).フ ラ ボ ノ イ ドの 抗酸 化活性 を 持 つ 構造 と し て B 環 の o 一ジ ヒ ド ロ オ キ シ 構造 , 2,3 位 が 2 重結 合 で ,4 位 が オ キ ソ 構 造, 3、5 位 に OH 基 が あ る こ と で あ る . 0 、 一が フ ラ ボ ノ ール と反応 す る こ とが 単離葉緑素細 胞 で 確 か め られ て い る .そ の 反応性 は 水酸基 の 位置や 数, 糖 の 結 合位 置 に よ っ て 変化 す る.パ ーオ キ シ.ダ ーゼ の 基 質 と して H 、02 を 除 去 す る こ と も ソ ラ マ メ,ム ラ サ キ ツ ユ ク サ の 孔 辺 細 胞 と表 皮 細 胞 で 報 告 で 行 わ れ る.OH ・の 除 去効 果 は ケ ル セ チ ン が最 も高 く,ケ ン フ ェ ロ ール , 3’一メ トキ シ ケ ル ゼ チ ン の 順 で あ り,3’が 配 糖体 に な っ て い る と活 性 が 強 く, OH 基 が 増す と 1徐.去機能 は 高 ま る .ま た ,脂質 の 過 酸化 反応 が 102 や OH ・に よ り開始 し,フ ラ ボ ノ イ ドは こ れ ら を 除 去 で き,脂 質 過 酸 化 反 応 を 抑制す る .過酸化脂 質 ラ ジ カ ル に よ る 白動 酸化 で も こ れ を除 く と予 想 され て い る.酸 化 さ れ た フ ラ ボ ノ イ ド は ア ス コ ル ビ ン 酸 に よ っ て 還 元 型 フ ラ ボ ノ イ ドに再 生 す る と考 え られ て い る (高濱 1988).フ ラ ボ ノ イ ドは お もに 表 皮細胞 に存 在 し,UV 光 の フ ィル タ ーと し て の 効 果 を示 し,ま た, UV に よ っ て 生合 成が 促進 さ れ る (浅 E日 2000)。 4. ア ス コ ル ビ ン 酸 ア ス コ ル ビ ン 酸 (AsA )は 02 −,OH ・や 102 を 消 去 す る が , H202 と の 反 応 は 生 体 で は 遅 い の で 無 視 で き る .さ ら に,Oz 一の 消 .去は SOD に よ る 消去 よ り遅 い .AsA に よ る OH ・の 消 去 は 早 い が,OH ・は 反 応 性 が 高 い た め , ’site − s.pecificoxidanti で あ るか ら,どの 程度 関与 す る か 不明 で あ る.ラ ジ カ ル の 除去 に も AsA は 関与 す る こ と が ダ イ ズ フ ォス フ ァ チ ジル コ リ ン の 酸 化 で 確か め られ て い る .こ の と き , α 一トコ フ ェ ロ ール と共 助 的 に 働 い て い る と考 え られ て い る (二 木 [988).AsA は抗 酸化機構 の 重要 な物 質 と して 後 述 の water −water サ イ ク ル や キ サ ン トフ ィ ル サ イ ク ル に 関 与 し,酸 化 α 一ト コ フ ェ ロ ール の 還 元 に 関 与 す る な ど の 役 害rlを 果 た し て い る (Alscher et al.1997 ). 5. α 一トコ フ ェ ロ ール 植 物 の 老 化 は老 化 導入 遺伝 子 に よ り発 現 で 誘 導 さ れ る. 光 に 関係 す る種 々 の 反応 で は活性 酸素 が 生 じ,膜 の 分解 が 引 き起 こ され る .過 酸化脂質 は 膜 の 選択 的膜 透過性 や 変性 を 起 こ す .し か し ,活 性酸 素 の 発 生 は老 化 の 後 期 に 生 じる た め 老化 の 引 き金 とは い え な い .また ,老 化過 程 で ミ ク ロ ボ デ ィ が プ ラス チ ッ ドに 密接 し, ミ ク ロ ボ デ ィ はペ ロ オ キ シ ソ ーム か らグ リオ キ シ ソ ーム 様微 小.器官 へ の 機能転 換 を 進 め ,脂質 の 移 動,脂肪 酸の β 酸 化や gluconogenesisへ の 利用 に お い て 重 要 な役割 を果 た す . トコ フ ェ ロ ール は 老 化 葉 に お け る こ れ らの 膜 機能 を維持す る 上 で 重 要で あ る . ト コ フ ェ ロ ール の 生 合 成 系 は 葉緑体 や 有色体 に あ っ て ,こ の 先 駆 物質 ’ホ モ ゲ ン チ ス . 酸 は 4一ヒ ド ロ オ キ シ フ ェ ニ ル ピ ル ビ ン 酸 か ら葉緑体 の ス トロ マ 側 に 局 在す る ジ オ キ シ ゲ ナ ー ゼ に よ り合 成 され る。葉緑体 の 老化 と同 様に 花 に も リポ オ キ シ ゲナ ーゼ が あ るた め, 花の 老 化 で 見 られ る膜 透過性 の 増 大 , サ イ トゾル の Ca の 増加,脂質 分解酵 素の 活性 化,リ ポ オ キ シ ゲ ナ ーゼ に よ る 脂 質か ら の 活性 酸 素 の 発 生 は起 こ る と 予想 さ れ る .葉 の 老化 中, α 一トコ フ ェ ロ ール は増加 し て ポ リ不 飽和 脂肪酸 と結 合 し, リポ オ キ シ ゲ ナ ーゼ を不活 性 化す る (Krupinska et a [.[998). 誌 IIL 活 1生酸素消去酵素 騫 鸛 L ス パ ーオ キシ ドヂ ス ム ターゼ (SOD ) SOD は 上 述 の よ う に 02一を 不 均 化 し (20 , 一† 2H +→ H ,O ,+ O ,),H / 02 を生 成 す る酵 素 で あ る. .酵 素 た ん ぱ く質 に 結 合す る金 属 イオ ン に よ り CuZnSOD ,MnSOD ,FeSOD の 3 種 類が 存在 す る.そ の 性 質 に つ い て は 表 1 に 示 した (浅 田 ら 1992)。CuZnSOD は 主 に 葉緑体 に 存在 し,サ イ トゾル に も あ る .菜 種 種 子 に は ペ ロ オ キ シ ソ ーム に も あ る . FeSOD は 葉 緑体,ミ トコ ン ド リア ,ペ ロ オ キ シ ソ ーム に 存 在 し,MnSOD は ミ トコ ン ド リア とペ ロ オ キ シ ソ ーム に 存 在 す る (Palma et al 、1998).カ ラ シ の 葉 で は FeSOD は 葉 緑体, CuZnSOD は ミ ト コ ン ド リア 膜 空間,MnSOD は ミ ト コ ン ド リア マ ト リ ッ ク ス に 分 布 し て い る (Salin 1988)。 SOD の 生 成誘 導 は 02 で あ り,E.coli の 場合 ,MnSOD と FeSOD の う ち, MnSOD の み が 誘 導 さ れ た が ,Sa (・char − omyces の 場 合 サ イ 】・ゾル の ZnCuSOD , ミ 1・コ ン ド リア MnSOD と も に 誘導 さ れ た (Fridovich 1974 ).西 洋 ク リの 葉 に は C.uZnSODI ,tl,llIの 3 つ の ア イ ソ ザ イム が あ り, 典 .型 的 な 老 化 の 症 状 が あ る 葉 で は そ れ ぞ れ 2.5.7 お よ び 4 倍 の 活 性 とな っ た (Longa et al.1994).02 ..が 組織 に 存在 す 表 1CuZnSOD ,MnSOD 、FeSOD 酵 素の 違 い 性 質 . .青緑色.酵母 ,植物,勤物 な か ら. 細胞質 ゾル ,リソ ソーム ,ミ トコ ン ドリ ア 内膜 ・外膜 の 空 間 に存在. 加熱,蛋 自分 解酵 素, 変 性剤に対 して 強 い 抵抗 を示 す . シ ア ン化合物は 強 い 阻 害剤.ジ エ チ ル ジチ オ カ ル バ ミ ン 酸 は 不活性 化す る . H 、O , に 長 時 間 さ ら され る と不 活性 化 す る. コ ム ギ種子 で は 2 つ の ア イ ソ ザ イ ム が あ る. M 冂 SOD 大腸 菌 か ら.桃色.上 記阻害剤で 阻害 れ る.熱,界面 活性 剤な どに 不安定 . 細菌,動物 ,植 物 か らも. ミ トコ ン ド リア マ ト リ ッ ク ス に存在. FleSOD 大腸 菌か ら.細 菌 ,藻類,植物 (43 属 3 属 に 分布.) IH202 に長 時間 さ らす と不活性化す る .一_一 一 一L−」二ニー一 一 一_−t』 .ア ジ化 物で 他の SOD 以 上 に 阻害 され 旨 る. る こ とは SOD の 活性 で 示 さ れ る.ま た ,H 、 O 、 に よ り不 活 性化 す る (河野 1988>.好 気性生物 は 高 い SOD お よび CAT 活性 を示.すが ,嫌気性 生物 の SOD 活性 は微 弱 で CAT 活性 を示 さ ない (Fridovich.1974 ). 2. ア ス コ ル ビ ン 酸 パ ーオ キ シ ダ ーゼ (APX ) APX は パ ーオ キ シ ダ ーゼ の 1 つ で ,葉緑体 チ ラ コ イ ド膜 表面 に存 在 し,H202 を無毒化 す る と と もに AsA を モ ノ デ ヒ ドロ ア ス コ ル ビ ン 酸 (MDA ) に 酸 化 す る (Elstner and Kramer tg73).し か し, エ ン ドウの 葉 で は ミ トコ ン ド リア とペ ロ オ キ シ ソ ーム に も存在 し,前者 の APX の グ ア イ ヤ コ ール と AsA に 対 す る 特性 は 類似 で あ る が ,後者 の APX は グ ア イ ヤ コ ール に 対 し て 高 い (JimEne∠ et al .tg98 ).こ の 酵素 は葉 緑体 SOD と と も に 光 合成 に お け る過 剰 e − , お よ び活性 酸 素 の 無毒 化 を図 り, 葉緑 体 の 機 能 を 維持 す る water −water サ イ ク ル を形成 す る (Asada 1999).酸化 AsA は グ ル タチ オ と,NADPH との 酸化還 元系 に よ り,AsA が 再 生 さ れ る (図 1,Alscher et al.1997). 3. 力 タ ラ ーゼ (CAT ) CAT は H 、0 、 を H20 と 02 に 分解 , 無.毒化す る.H ,,O 、 の 濃度 が 低 い と き はパ ーオ キ シ ダ ーゼ 活性 を示 し,RH , + H2 02→ R + 2H、 0 (RH2 還 元 物 質)の 反 応 に よ り,還元物質 を 酸 化 し, H 、0 , 濃 度が 高 い と き は 2H ,0 、 →2H 、〇 + O 、 の 反 応 に よ 1),酸 素 を発生 す る. 細 胞 内 の 存 在 部位 は ト ウ モ ロ コ シ に お い て は サ イ ト ゾ ル ,グ リオ キ シ ソ ーム ,ペ ロ オ キ シ ソ ーム , ミ トコ ン ド リ ア に 存 在す る が ,一般 に 葉緑体,ミ トコ ン ド リア な どで 生 成 した H , 0 , は ペ ロ オ キ シ ソ ーム に 移 行 し , 大 部分 は こ こ の CAT で 無毒化 さ れ る .前述 の トウ モ ロ コ シ で は 3 ア イ ソザ イ ム が 知 られ , 表 2 に 示 す 性質 を 持 っ て い る (Scan一dalios1994).CAT は 02一に よっ て 阻 害 さ れ る (Kono and Fridovich l982). S、 IV .環境 と 活性酸 素 髏蠢 1. 活 性酸素の 生 成 と環境要 因 植 物細胞 で は活性酸 素 は通常 に 発生 して い る が , 種 々 の ス ト レ ス が加 わ る と発 生 は 助長 され る.ス トレ ス の 生 物 的 要 因 と し て 病 原菌 (.毒素),傷 (害虫 な どに よ る ),養分 欠 乏 症 や 過 剰 症 な ど が あ り,非生 物的要 因 と し て 温 度 (高温 , 低 温 ),光 (強 光 ,弱 光 ),UV (Baumbusch et u ].1998 ; Ranieri et al .1996 ), 湛水 (Ushimaru et al.1992 ), 乾燥, 除草剤 (KnOrzer et al.1996),オ ゾ ン (Baumbusch et al . 1998),SO2,無機 イ オ ン ,高塩類 な どが あ る .ま た ,ス ト レ ス 要因 が 重な る と よ り一層 ス トレ ス が 強 くな り,活性 酸 素 の 発 生 や そ の 消去 酵素 の 活性 変動 は著 し くな る (Alsche1・ et ai .1997). 2. 活 性酸素 の 生 成 と消去機構 1) 正 常→ス トレ ス →順化 の 過程 生体 は正 常 な状態 で も活性 酸素 を生 ず る た め ,こ れ を除 去 す る機構 が 備 っ て い る.葉 緑体 で は e ..が 酸素 を還元 し て , 02 一を生 じ, O , 一を SOD に よ り H , 0 、 に 変 え ,APX に よ り分解 され る .ペ ロ オ キ シ ソーム で も, ミ トコ ン ドリ ア で も常時 活性 酸 素 を 発 生 し,こ れ ら は 活 性 酸 素消 去 酵 素 (active oxygen species =AOS scavenging enzyme ) に よ っ て 無毒化 され る.ス トレ ス が 生 じた 場 合,正 常な 代謝 や 生 理 反 応 は 進 行 し な くな り, 活 性 酸 素 の 発 生 を 助 .長 す る (Alscherand Hess 且993).ス ト レ ス 種 が 消 去 され ず ,活 性 酸素 も残 る と DNA に 影 響 を与 え,つ い で t −RNA に 影 響 し ,AOS scavenging enzynle の 活性増加 や 抗 酸化物質 の 含 量変化 な ど を と も な っ て 活性 酸素 を 消.去す る .ス ト レ ス 自 体 が な くな れ ば, あ る い は ス トレ ス に 適応 (順 化 )す れ ば, 植 物体 は 元 の 状態 に 回 復 , あ る い は適 応 した状 態 に な る. 2> 葉緑 体 に お け る 3 つ の 抗 酸化機構 water water サ イ ク ル (Asada [999) また は Mehle1・APX 一 光 呼 吸 (Osmond and Grace I995) 強 光 ,高温 お よ び 低 CO ,,な ど の 条件 で は 葉 緑体 の PSIl に お け る H 、0 の 分 解 物 で あ る e −,お よ び PSI で .生 成 す る NADPH は過 剰 と な る.e一は 02 と 反 応 し ,02一を 作 り, SOD で さ ら に H202 に 変 え, さ ら に H ,O , を APX に よ り AsA を基 質 と し て 比 0 と モ ノ デ ヒ ドロ ア ス コ ル ビ ン 酸 (MDA )に 変 え,無 毒化 す る .こ の 反 応で は AsA が 減 少す るの で ,MDA を グル タ チ オ ン (GSH )で 還元 し,AsA を 再 生 す る.酸化 GSH (GSSG )は過剰 とな っ て い る NADPH を使 っ て ,GSH を再 生 す る (AsA プ ール 〉.つ ま り, PSI に お け る H ,0 の 光 分 解 に 始 っ た 過 剰 な e一と PSII に お け る 過剰 な NADPH を H 、 0 に もどす こ と で ,余分 な 光 エ ネル ギ ーの 堰 とな っ て い る (図 1,Alscher et al .1997). キサ ン トフ ィ ル サ イ ク ル も う一っ の 過剰 な 光 を 分 散 さ せ る機 構 と し て 存在 す る. ビ オ ラ キ サ ン チ ン は デ エ ポ キ シ ゲ ナ ーゼ に よ リア ン テ ラ キ サ ン チ ン を 経 て ゼ ア キ サ ン チ ン に 変 換 さ れ る .こ の と き AsA が 使 わ れ る が , 酸化 AsA は PSI の フ ェ デ ロ キ シ ン に よ っ て AsA に 再生 さ れ る.ゼ ア キサ ン チ ン は夜 , エ ポ キ シ ダ ーゼで ア ン テ ラ キ サ ン チ ン を 経て ビ オ ラ キサ ン チ ン に 酸 化 さ れ る (図 1,A ]scher et aL ]997,Demmig −Adams and Adams 1993).多 くの 植 物 で は 陽光植物 は 陰性植物 よ り こ の サ イ ク ル の 色 素含 量 が 高 く, .両者 の 差異 は C4 植物 よ り C3 植物 で よ り大 き くな る (Brugnoliet a [,1998). ク ロ ロ フ ィ ル 分解系 強光 に よ る 光エ ネ ル ギ ー.過 剰 は葉緑体 を減少 す る こ と に よ っ て も達 せ られ る .こ れ に は ク ロ ロ フ ィ ラ ーゼ ,ク ロ ロ フ ィ ル オ キ シ ダーゼ や ク W ロ フ ィ ル パ ーオ キ シ ダ ーゼ が 関 与 す る可 能 性 が あ る.ク ロ ロ フ ィ ル 分 解 の in 1,iVO反 応 系 に 関 す る研 究 は 老 化 葉 や エ チ レ ン 処 理 値 物 体 で 研 究 され て い る が (Abe [es et a1,1988 ; Huff 1982 ; Shimekawa et al . 1978 ; Yamauchi and Minamide 1985),葉緑体 の 抗酸化機 能 の 面か ら の 検討 は 少 な い .今後 の 研 究課題 で あ ろ う. 強光 に よ り葉緑素 含 量は低 下す る.上記以外 に 直接的 に 02 一や OH ・が 葉緑 素 を分 解 して ,強 光 に 対 抗 して い る一 方 ,葉 緑 体 が 細 胞 内 で 配 列 を 変 え,対 抗 も して い る (林 2000). ポ リフ ェ ノ ール オ キ シ ダー一ゼ (PPO ) PPO は プ ラ ス チ ッ .ドに 存在 し,葉緑体 に お い て は チ ラ コ イ ド膜 上 の PS [に 存 在 し, lightharvestingpigment に結 合 して い る (Liebereiand Biehl 1976).PPO は C 、植物 の 維 管 朿鞘細胞 で は 後生 的 に 欠 き ,孔辺 細胞 の プ ラ ス チ ッ ドで も 欠 く (Vaugha et a1 .1988).PPO は フ ェ ノ ール 性化合物 の 水酸化 (生合 成的 に は 関与せ ず ,傷.害 に よ る 酸 化 に 関与 ), キ ノ ン 生成 と重 合, 病害抵 抗性 に 関与 し, ま た, 且AA との 関連 で 発 根 に 関 与 す る と考 え られ て い る.さ ら に Vaughu et al .(1988)は PPO が PSI に存在 す る こ とか ら葉 緑体 の エ ネ ル ギ ー変 換 の 役割 が あ る と予 想 して い る .つ ま り,02 レ ベ ル を 一定 水 準 に 保 っ て Mehler 反 応 を制 御 す る.模式 的 に ポ リフ ェ ノ ール 性 物 質 は PPO に よ っ て キ ノ ン に 酸 化 され , キ ノ ン は PS ]で e一に よ っ て ポ .リ フ ェ ノ ール 性 物 質 に 再生 さ れ る .
V . 活性酸素 消去酵 素 と植物生 長調節物 質 臨 活 性酸勲 溌 生 お よ び消去 と 植物生 長 講 肋 質 に 関 連 す る記 載 や報 告 は少 な く,今後 の 研究 が必 要 な と こ ろ で あ る .オ ーキ シ ン 関 連 で は トウ モ ロ コ シ の 発 芽時 に お け る オ ーキ シ ン に よ る CAT 遺 伝 了 発 現 が 調 査 さ れ ,オ ーキ シ ン に よ り 3 種 の 未熟胚 中の CAT 遺伝 子 お よ び酵 素 が 誘 導 さ れ , 全 CAT 活性 は オ ーキ シ ン 濃度 に 対応 し た.3 種類 の CAT 遺 伝 子 の 転写 は オ ーキ シ ン 処 理 後 の 時間 に よ っ て 異 な り,オ ーキ シ ン 処 理 に よ る 未熟胚 中の 変化 は 発芽 種子 に お け る も の と 似 た (Guan and Scandalios.2002 )。また ,ABA に つ い て は 10−4mol の ABA 添 加 培 地 で は 受 粉 後 18−21 日の トウ モ ロ コ シ の 胚 は 発芽 せ ず ,成熟 を続 け る の に 対 し, シ ュ ク ロ ーズ /ミネ ラ ル 培地 で 発芽 す る.ABA 添加培 地で は胚 盤 の 蛋 白 は CAT .1 お よ び CAT −3 蛋 白 が 集 積 す る (CAT ..2 は しな い )が ,発育 す る胚 で は CAT −2 蛋 自 は集積 す る が ,CAT −1 は 減少 す る .CAT −2 の 発現 は ABA の ダウ ン レ ギ ュ レ イ トを 受 け ,CAT 1 と CAT −3 は 正 の ABA 制 御 を 受 け る候補者 と 言 え そ うで ある .胚盤 RNA の North − ern −analysis で は Cat1RNA は ABA の 存 在 .ドで 増加 し,無 け れ ば 増 加 せ ず . , (/at2RNA は ABA 無 添 加 で 増 加 す る. CA T3 は 外生的 ABA に は反 応 し な い 結果 で あ っ た (Scan− dulios1994).10 あ る い は 100 μ molABA を トウ モ ロ コ シ に 処理 す る と葉 の 02 一と H202 レ ベ ル は高 ま り,SOD 、APX , CAT な ど の 活性 も 高 くな る (Jiang and Zhang 200D . 抗 酸化酵素 と エ チ レ ン と の 関係 は成 熟 あ る い は 老 化 関 連 で 調 べ られ て い る.リ ン ゴ 成熟 の 開始 を引 き 起 こ す エ チ レ ン の 発 生 は SOD と CAT と.関係 が あ り, エ チ レ ン の ク リマ ク テ リ ッ ク ピ ーク は SOD と CAT 活 性 ピーク と 重 な る (Mesia 1998).また , ホ ウ レ ン ソ ウ の 低 温耐性 に は ポ リア ミ ン が 関 与 し, 低 温 処 理 中 S ア デ ノ シ ル メ チ オ ニ ン (SAM )脱 炭酸 酵 素 は ス ペ ル ミ ジ ン の 増加 と比 例 して 活 性 増 加 す る .こ の 物 質 は 光 合成 装置 の 保 護 に 関与 して い る (He et al .2002)が ,SAM は エ チ レ ン 生合成 の 先駆物 質で あ る の で , エ チ レ ン 生成 と 関係が あ る か も しれ な い .同 様 に , 葉 を乾燥 した と き の ス トレ ス 性 エ チ レ ン生 成 は ア ミ ノ シ ク ロ プ ロ パ ン カ ル ボ ン 酸 (ACC )や メ チ ル 化 ACC の 集 積 と 関係 し,こ の 生 成 は ACC 合成抑 制剤 (AOA )や フ リー ラ ジ カ ル 消去 剤 (没食子酸 n 一プ ロ ピル )で抑 制 さ れ る.後 者 は ACC に は 影 響 せ ず,メ チ ル 化 ACC を増 加 させ , リポ キ シ ゲ ナ ーゼ 活 性 を 高め な い の で ,フ リーラ ジ カ ル を増 加 さ せ な い (Kacperska and Zebalska 1989).




